Neuroscience Tutorial An illustrated guide to the essential basics of clinical neuroscience
Found at: http://thalamus.wustl.edu/course/
SOMATOSENSORY PATHWAYS FROM THE BODY
A. Peripheral receptors:
There are three different categories (modalities) of the somatosensory system. The first, discriminative touch, is the perception of pressure, vibration, and texture. This system relies on four different receptors in the skin. They are:
1) Meissner's corpuscles 2) Pacinian corpuscles 3) Merkel's disks 4) Ruffini endings
The first two are considered rapidly adapting (they quickly stop firing in response to a constant stimulus) and the second two are considered slowly adapting (they do not stop firing). To put this into an example, if you lay your pen down in your palm, the Meissner's and Pacinian corpuscles will fire rapidly as it first touches down, to let you know something has landed. If the pen lays still, they will stop firing almost right away. The Merkel's and Ruffini endings, however, will continue to fire to let you know that something is still there.
The pain and temperature system does not have specialized receptor organs. Instead, it uses free nerve endings throughout skin, muscle, bone, and connective tissue to perceive changes in temperature and pain peptides. Although pain will result from damage to a free nerve ending, in reality most pain is a result of substances released by damaged tissues: prostaglandins, histamine, and substance P. The free nerve ending has receptors for these substances and lets you know (stridently) when tissue has been damaged.
The third modality, proprioceptive sensation, relies on receptors in muscles and joints. The muscle spindle is the major stretch receptor within muscles, and just like the cutaneous receptors, it has a rapidly-adapting and slowly-adapting component. (For more on the muscle spindle see "Spinal motor structures".) There are also Golgi tendon organs and joint afferents to monitor stresses and forces at the tendons and joints.
B. Axon diameters:
Sensory axons can be classified according to diameter and therefore conduction velocity. The largest and fastest axons are called Aa, and include some of the proprioceptive neurons, such as the stretch receptor. (Note: there is a separate, Roman numeral classification used for the proprioceptive axons, which will be covered in the section on muscle spindles.) The second largest group is called Ab, which includes all of the discriminative touch receptors. Pain and temperature include the third and fourth groups, Ad and C fibers. There are two subtypes of pain. "Fast pain", carried by the Ad fibers, is the instantaneous pain that makes your arm jerk back before you even realized you were burned. It is sharp and piercing and over quickly. "Slow pain" is carried by C fibers. C fibers are not only small, they are unmyelinated (the only sensory axons without myelin), so their conduction velocity is quite slow. Slow pain is primarily mediated by those tissue-damage peptides listed above, and can go on indefinitely. It is distressing, it can be dull and aching, and it does not trigger withdrawal reflexes like the fast pain. A perfect example of slow pain is when you stub your toe on the coffee table. You feel the jolt of impact (proprioception and Pacinian corpuscles), and you have approximately a heartbeat to think, "This is really going to hurt." That heartbeat is the C-fiber travel time from your toe to your brain. When the signal hits, the pain is severe and lasts for quite a while. It is, however, a nice demonstration of the relative conduction velocities of Aa and C fibers.
As the dorsal root enters the cord (all sensory information comes in via the dorsal root, and all sensory cell bodies are in the dorsal root ganglion), the fibers sort themselves out by diameter. The largest fibers enter the cord most medially, and the smallest fibers enter most laterally. From there, the three modalities take very different paths, so we must look at each one separately.
C. The discriminative touch system:
The posterior columns should be a review from the "Basic somatosensory" section. Here is a schematic of the pathway, to remind you:

The key points are that the primary afferents ascend all the way to the medulla, on the ispilateral side of the cord, in the posterior columns. The secondary afferents cross in the medulla and ascend as the medial lemniscus. In the thalamus they synapse in the VPL (the ventroposterior lateral nucleus) and finally ascend to cortex.
D. The pain and temperature system:
This system shares one major rule with the discriminative touch system: primary afferents synapse ipsilaterally, then secondary afferents cross. SYNAPSE, then CROSS. The crossings just occur at different levels.
Pain afferents (all of the following applies to temperature as well) enter the cord laterally, due to their small size, and synapse more or less immediately. "More or less", because they actually can travel one or two segments up or down the cord before synapsing. Lissauer's tract is the tract carrying these migrating axons, but they are only in the tract for a short time. Within one or two levels, they enter the dorsal horn and synapse.

The dorsal horn is a multi-layered structure. The thin outermost layer is called the posterior marginalis layer. The wide pale second layer is called the substantia gelatinosa, and the layer deep to that is called the nucleus proprius. The layers continue into the ventral horn, but these are the three significant ones for now.

The two types of pain fibers enter different layers of the dorsal horn. Ad fibers enter the posterior marginalis and the nucleus proprius, and synapse on a second set of neurons. These are the secondary afferents (purple, below) which will carry the signal to the thalamus. The secondary afferents from both layers cross to the opposite side of the spinal cord and ascend in a tract called (logically) the spinothalamic tract. Tracts are always labeled from beginning to end.
The C fibers enter the substantia gelatinosa and synapse, but they do not synapse on secondary afferents. Instead they synapse on interneurons - neurons which do not project out of the immediate area. The interneurons must carry the signal to the secondary afferents in either the posterior marginalis or the nucleus proprius.
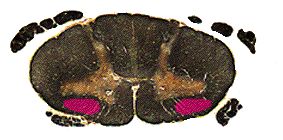
The spinothalamic tract ascends the entire length of the cord as shown above, and the entire brainstem, staying in about the same location all the way up. Below are representative slides showing the tract in the medulla and midbrain. Notice that by midbrain the spinothalamic tract appears to be continuous with the medial lemniscus. They will enter the VPL of the thalamus together. 
The spinothalamic system enters the VPL, synapses, and is finally carried to cortex by the thalamocortical neurons. Here is a schematic of the entire pathway:
E. Pain control:
It has been recognized for centuries that opium and related compounds (such as morphine) are powerful analgesics. Several decades ago scientists hunted down the opiate receptor which was responsible for the potent effects. They then reasoned that if there was such a receptor in the body, maybe the body used its own endogenous form of opium to control pain. (It has also been recognized for centuries that under certain circumstances, i.e. the heat of battle, a serious wound may not cause pain.) This hypothetical compound was named "endorphin", from endogenous-morphine. Soon after, an entire class of peptide neurotransmitters was discovered that interacted with the opiate receptor, and now includes endorphins, enkephalins, and dynorphins. Synthetic, exogenous forms of these compounds continue to be discovered, prescribed, and abused, and are classed under the general term, "narcotics".
There are opiate receptors throughout the central nervous system. In the dorsal horn, they are located on the terminals of the primary afferents, as well as on the cell bodies of the secondary afferents. Opiate interneurons in the spinal cord can be activated by descending projections from the brainstem (especially the raphe nuclei and periaqueductal grey), and can block pain transmission at two sites. 1) They can prevent the primary afferent from passing on its signal by blocking neurotransmitter release, and 2) they can inhibit the secondary afferent so it does not send the signal up the spinothalamic tract.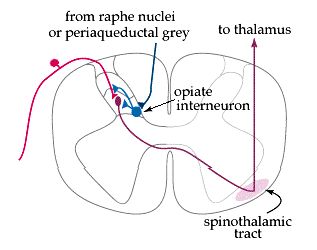
F. The proprioceptive system:
The proprioceptive system arises from primarily the Aa afferents entering the spinal cord. These are the afferents from muscle spindles, Golgi tendon organs, and joint receptors. The axons travel for a little while with the discriminative touch system, in the posterior columns. Within a few segments, however, the proprioceptive information slips out of the dorsal white matter and synapses. After synapsing it ascends without crossing to the cerebellum.
Exactly where the axons synapse depends upon whether they originated in the legs or the arms. Leg fibers enter the cord at sacral or lumbar levels, ascend to the upper lumbar segments, and synapse in a medial nucleus called Clarke's nucleus (or nucleus dorsalis). The secondary afferents then enter the dorsal spinocerebellar tract on the lateral edge of the cord.
Fibers from the arm enter at cervical levels and ascend to the caudal medulla. Once there they synapse in a nucleus called the external cuneate (or lateral cuneate) nucleus, and the secondary axons join the leg information in the dorsal spinocerebellar tract. 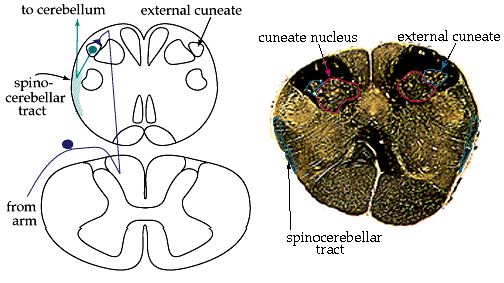
The spinocerebellar tract stays on the lateral margin of the brainstem all the way up the medulla. Just before reaching the pons, it is joined by a large projection from the inferior olive. These axons together make up the bulk of the inferior cerebellar peduncle, which grows right out of the lateral medulla and enters the cerebellum.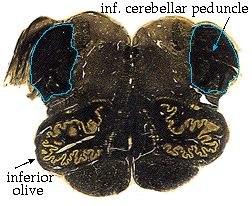
The figures above outline the course of the dorsal spinocerebellar tract. Surely, there must be a ventral spinocerebellar tract? Naturally, there is, and it travels in approximately the same place - the lateral margin of the spinal cord, just ventral to the dorsal spinocerebellar tract. The two cannot be distinguished in a normal myelin stain. The ventral spinocerebellar tract seems to defy the ipsilaterality of the cerebellum, because the fibers entering it in the spinal cord actually cross on their way into the tract. However, they (somewhat inefficiently) cross back before entering the cerebellum. Therefore the cerebellum still gets information from the ipsilateral body.
A note about generalizations:
There is actually a fair amount of mixing that goes on between the tracts. Some light touch information travels in the spinothalamic tract, so that lesioning the dorsal columns will not completely knock out touch and pressure sensation. Some proprioception also travels in the dorsal columns, and follows the medial lemniscus all the way to the cortex, so there is conscious awareness of body position and movement. The pain and temperature system, although it does ascend to somatosensory cortex, also has multiple targets in the brainstem and other areas.
For a more interactive tutorial on the anatomy, and for stunning three dimensional pictures of these pathways, try the Digital Anatomist in the "Other links" section.